|
تضامنًا مع حق الشعب الفلسطيني |
باحة برودمان العاشرة
| باحة برودمان العاشرة | |
|---|---|
| الاسم العلمي Area frontopolaris |
|
 | |
 | |
| تفاصيل | |
| نيوروليكس | Brodmann area 10 |
| تعديل مصدري - تعديل | |
باحة برودمان العاشرة (BA10، القطب الأمامي للقشر أمام الجبهي، على الوجه الجانبي للقشر أمام الجبهي، أو القشر أمام الجبهي الأمامي) هي الجزء الأكثر أمامية من القشر أمام الجبهي في دماغ الإنسان.[1] بصورة عامة، هذه الباحة عُينت أصلاً بلغة التهندس الخلوي الخاصة بها كما لوحظ في أدمغة الجثث، لكن وبما أن التصوير الوظيفي الحديث لا يمكنه تحديد هذه الحواف بدقة، فإن مصطلحات قشر أمام الجبهي الأمامي، قشر أمام الجبهي الجانبي والقطب الأمامي للقشر أمام الجبهي تُستخدم لتدل على المنطقة الأكثر أمامية من القشر الجبهي التي تغطي باحة برودمان العاشرة تقريباًـــ ببساطة، فإن هذا يؤكد حقيقة أن باحة برودمان العاشرة لا تتضمن كل أجزاء القشر أمام الجبهي. باحة برودمان العاشرة BA10 هي المنطقة الهندسية الخلوية الأكبر في دماغ الإنسان. ووصفت بأنها «واحدة أقل المناطق التي فُهمت جيداً في دماغ الإنسان».[2] الأبحاث الحالية تقترح مساهمتها في العمليات الإستراتيجية لاستعادة الذاكرة ومختلف الوظائف التنفيذية. يفترض أن وظائف هذه المنطقة قد نتجت عن اتساعها بالنسبة لبقية الدماغ خلال تطور الإنسان.[3]
التشريح
الحجم
يبلغ حجم باحة برودمان العاشرة حوالي 14 سم3 وتشكل تقريبا 1-2% من كامل حجم الدماغ. بالمقارنة، فإن حجم باحة برودمان العاشرة عند قرود البونوبو (البابون) تبلغ حوالي 2.8 سم3، وتشكل فقط 0.74% من حجم دماغها. في كل نصف كرة مخية، تحوي الباحة العاشرة ما يقدر بـ 250 مليون عصبون.[3]
الموقع
باحة برودمان العاشرة هي قسم من المنطقة الجبهية محددة التهندس الخلوي من القشرة المخية. تشغل الأجزاء الأكثر جانبية (منقارية) من التلفيف الجبهي العلوي والتلفيف الجبهي المتوسط. عند البشر، تكون محددة من الأسفل بالثلم المنقاري العلوي في الجانب الإنسي من نصف الكرة المخية. ولا تتمدد على طول الثلم الحزامي. وتكون محددة بالتهندس الخلوي من الأعلى بالباحة 9 الجبهية الحبيبية، ومن الخلف بالباحة 46 الجبهية المتوسطة، ومن الأسفل بالباحة 47 الحجاجية والباحة 12 المنقارية أو، بحسب الرواية القديمة للخريطة القشرية لبرودمان (برودمان- 1909)، باحة برودمان 11 الأمامية الجبهية- 1909.[4] تتوضع الباحة 10 في أعلى الجيب الهوائي العظمي الذي تحددت البحوث الفيزيولوجية الكهربية عليه.[5]
العلاقة مع القطب الجبهي
منطقة القطب الجبهي من القشرة أمام الجبهية -عند البشر- لا تتضمن فقط الباحة 10 بل هناك جزء من الباحة 9. باحة برودمان 10 تمتد خلف المنطقة القطبية باتجاه جانبها الإنسي السفلي. عند القرد (هجرس أو جينون)، المنطقة القطبية ممتلئة بباحة برودمان 12 (باحة برودمان 10 عنده وجدت في المنطقة أمام جبهية الحجاجية).[2]
البنية الخلوية
عند البشر، وصفت الطبقات القشرية الست من الباحة 10 بأنها تملك «مظهراً متجانساً ملحوظاً».[3] جميعها يمكن أن تحدد بسهولة. بالنسبة لبعضها، الطبقة I رقيقة إلى متوسطة العرض مشكلة 11% من عمق الباحة 10. الباحة II رقيقة وتحوي خلايا حبيبة صغيرة وهرمية متوسطة إلى داكنة التلوين (بطريقة تلوين نيسل) التي تلون الـRNA و الـDNA. الطبقة الأعرض هي الطبقة III. عصبوناتها الهرمية أصغر وأقرب إلى الطبقة العلوية II من الطبقة السفلية IV. ومثل الطبقة II تكون خلاياها متوسطة إلى داكنة. الطبقات II و III تشكل 43% من عمق القشر. الطبقة IV لها حدود واضحة مع الطبقة III أعلاها والطبقة V في الأسفل، وهي رقيقة. خلاياها شاحبة إلى متوسطة التلوين. الطبقة V عريضة وتحتوي طبقتين فرعيتين، Va و Vb. كثافة خلايا Va أكبر من Vb وتلوينها داكن أكثر. الطبقات IV و V تشكل 40% من الكثافة القشرية. الطبقة VI تحت الطبقة V وفوق المادة البيضاء تحوي عصبونات داكنة هرمية ومغزلية الشكل، وهي تسهم في تشكيل 6% من سماكة الباحة 10.[3] تختلف الباحة 10 عن مجاورتها الباحة 9 بأن الأخيرة لها طبقة Vb مميزة أكثر والطبقة II أكثر وضوحاً. الباحة 11 المجاورة مقارنةً مع الباحة 10 لها طبقة IV أرق وطبقات Va و Vb و II أكثر وضوحاً.[3] تملك الباحة 10 عند البشر الكثافة العصبونية الأقل بين أدمغة الرئيسيات.[3] وهي أيضاً استثنائية في كون عصبوناتها تملك تشجراً تغصنياً واسعاً وكثافة عالية مع نتوءات تغصنية.[6] فسرت هذه الحالة بأنها تحقق التكامل بين المدخَلات من مختلف الباحات.[2]
الباحات الفرعية
تقسم الباحة 10 إلى ثلاث باحات فرعية، 10p، 10m و10r ا. 10p تشغل القطب الجبهي بينما تغطي الباقيتان الجزء الإنسي البطني من القشر أمام الجبهي.[7] الباحة 10m لها طبقات II و IV رقيقة وطبقة V أكثر وضوحاً. بالمقابل، 10r تملك طبقة II أوضح وطبقة V أثخن. الخلايا الهرمية الكبيرة توجد أيضاً في 10r الطبقة III وبدرجة أكبر في 10p. لكن لوحظ أن «الاختلافات بين الباحات الثلاث تدريجية، ومن الصعب تمييز الحدود بدقة فيما بينها».[7]
الارتباطات
الأبحاث المجراة على الثدييات (الرئيسيات) تقترح كون الباحة 10 تملك ارتباطات إدخال وإخراج مع باحات عمليات الربط العليا القشرية الأخرى، بشكل خاص في القشر أمام الجبهي، بينما تملك ارتباطات أقل مع الباحات الحركية أو الحسية الأولية. ارتباطاتها عبر المحفظة القصوى تصلها مع الباحات السمعية ومتعددة الحواس للثلم الصدغي العلوي. وتستمر أيضاً في الحزمة الطولانية الإنسية في المادة البيضاء لباحات التلفيف الصدغي العلوي على التلفيف الصدغي العلوي (الباحات TAa، TS2، و TS3) والباحات متعددة الحواس المجاورة في البنك العلوي للثلم الصدغي العلوي (TPO). هناك باحة أخرى مرتبطة عبر المحفظة القصوى هي المنطقة البطنية للجزيرة. الارتباطات عبر القشرة الحزامية تصل الباحة 10 مع القشر الحزامي الأمامي الخلفي، والقشر خلف الطحالي. الحزمة الشصية تربطها مع اللوزة، القشر السوي proisocortex القطبي الصدغي والجزء الأمامي من التلفيف الصدغي العلوي. لا توجد ارتباطات مع القشر الجداري، القشر القذالي ولا القشر الصدغي السفلي.[8] لخصت ارتباطاتها بأنها " تبدو غير متصلة مع “مجرى” الباحات الذي تسلكه الباحات أمام الجبهية الأخرى. هي المنطقة أمام الجبهية الوحيدة المترابطة بشكل مسيطر (وربما على وجه خاص) مع القشر Supramodal في الـPFC، القشر الصدغي الأمامي والقشر الحزامي.[9] هناك اقتراح هو أنه وبسبب ارتباطاتها هذه فإنه يمكن لها أن "تلعب دوراً كبيراً في المستويات العليا من التكامل والدمج بين المعلومات القادمة من أجهزة البصر، السمع والحس الجسمي لتنجز التفسير المفهوم التجريدي الشكلي للبيئة. ويمكن أن يكون الأساس التشريحي للدور المقترَح للقشر أمام الجبهي المنقاري في التأثير في معالجة المعلومات التجريدية والدمج والربط بين نتائج العمليات المعرفية العديدة".[8]
التطور
افترضت كاترينا سيميندفيري Katerina Semendeferi وزملاؤها أنه «خلال تطور أسلاف الإنسان»، خضعت الباحة 10 لتغيرين: أحدهما يتضمن زيادة معتبرة في الحجم الكلي، والآخر يتضمن زيادة خاصة في الارتباط، بشكل خاص مع باحات تتصف بعمليات ربط عليا أخرى.[3] القوالب الداخلية القحفية المأخوذة من داخل جمجمة إنسان فلوريس أظهرت توسعاً في منطقة القطب الجبهي بما يفترض تضخماً في باحة برودمان 10.[10]
الوظيفة
بالرغم من اتساع هذه المنطقة عند الإنسان، إلا أن وظيفتها لم تفهم بشكل جيد.[3] اقترح كاتشلين وهايافل Koechlin & Hyafil أن معالجة «التشعب المعرفي» هي لب وظيفة القشرة أمام الجبهية.[11] التشعب المعرفي يمكّن المهمة السابقة لتكون مصونة في حالة انتظار للاسترجاع فيما بعد والتنفيذ بُعيد إتمام المهمة الجارية. العديد من سلوكياتنا المعقدة ونشاطاتنا الذهنية تتطلب اندماجا متزامنا للعديد من المهام في آن واحد، اقتُرح أنه ربما تكون القشرة أمام جبهية الأمامية هي التي تؤدي الوظيفة بمجال شامل في هذه العمليات المجدولة. وهكذا، تشارك القشرة أمام جبهية المميزات مع الإدارة المركزية في نموذج باديلي Baddeley's model للذاكرة العاملة. إلا أن هناك فرضيات أخرى قُدمت أيضاً، مثل التي قدمها Burgess et al .[12] هذه الفرضيات تأخذ أيضاً تأثير الجهاز اللمبي بعين الاعتبار، حيث القشر القطبي الجبهي متصل عبر القشر أمام الجبهي الإنسي البطني. التحليل التجميعي لـ2006 وجد أن القشر أمام الجبهي المنقاري مشارك في الذاكرة العاملة، الذاكرة النوبية، والتنسيق متعدد المهام.[13]
صور
-
 صورة متحركة.
صورة متحركة. -
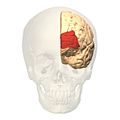 منظر أمامي.
منظر أمامي. -
 منظر وحشي.
منظر وحشي. -
 منظر أنسي.
منظر أنسي.




انظر أيضًا
| في كومنز صور وملفات عن: باحة برودمان العاشرة |
مراجع
- ^ Knowlton، Barbara J.؛ Morrison, Robert G.؛ Hummel, John E.؛ Holyoak, Keith J. (يوليو 2012). "A neurocomputational system for relational reasoning". Trends in Cognitive Sciences. ج. 16 ع. 7: 373–381. DOI:10.1016/j.tics.2012.06.002.
- ^ أ ب ت Ramnani N, Owen AM. (2004). Anterior prefrontal cortex: insights into function from anatomy and neuroimaging. Nat Rev Neurosci. 5(3):184-94. دُوِي:10.1038/nrn1343 ببمد: 14976518
- ^ أ ب ت ث ج ح خ د Semendeferi K, Armstrong E, Schleicher A, Zilles K, Van Hoesen GW. (2001). Prefrontal cortex in humans and apes: a comparative study of area 10. Am J Phys Anthropol. 114(3):224-41. دُوِي:10.1002/ajpa.20947 ببمد: 11241188
- ^
 تتضمن هذه المقالة text متاحًا تحت رُخصة CC BY 3.0. "نسخة مؤرشفة". مؤرشف من الأصل في 2013-12-10. اطلع عليه بتاريخ 2016-11-09.
تتضمن هذه المقالة text متاحًا تحت رُخصة CC BY 3.0. "نسخة مؤرشفة". مؤرشف من الأصل في 2013-12-10. اطلع عليه بتاريخ 2016-11-09.{{استشهاد ويب}}: صيانة الاستشهاد: BOT: original URL status unknown (link) - ^ Wallis JD. (2010). Polar exploration. Nat Neurosci. 13(1):7-8. دُوِي:10.1038/nn0110-7 ببمد: 20033080
- ^ Jacobs B, Schall M, Prather M, Kapler E, Driscoll L, Baca S, Jacobs J, Ford K, Wainwright M, Treml M. (2001). Regional dendritic and spine variation in human cerebral cortex: a quantitative golgi study. Cereb Cortex. 11(6):558-71. دُوِي:10.1093/cercor/11.6.558 ببمد: 11375917 نسخة محفوظة 03 مارس 2006 على موقع واي باك مشين.
- ^ أ ب Ongür D, Ferry AT, Price JL. (2003). Architectonic subdivision of the human orbital and medial prefrontal cortex. J Comp Neurol. 460(3):425-49. دُوِي:10.1002/cne.10609 ببمد: 12692859
- ^ أ ب Petrides M, Pandya DN. (2007). Efferent association pathways from the rostral prefrontal cortex in the macaque monkey. J Neurosci. 27(43):11573-86. دُوِي:10.1523/JNEUROSCI.2419-07.2007 ببمد: 17959800 "نسخة مؤرشفة". مؤرشف من الأصل في 2008-11-22. اطلع عليه بتاريخ 2016-11-09.
{{استشهاد ويب}}: صيانة الاستشهاد: BOT: original URL status unknown (link) - ^ Ramnani N, Owen AM. (2004). Anterior prefrontal cortex: insights into function from anatomy and neuroimaging. Nat Rev Neurosci. Mar;5(3):184-94. دُوِي:10.1038/nrn1343 ببمد: 14976518
- ^ Falk D, Hildebolt C, Smith K, Morwood MJ, Sutikna T, Brown P, Jatmiko, Saptomo EW, Brunsden B, Prior F. (2005). The brain of LB1, Homo floresiensis. Science. 308(5719):242-5. دُوِي:10.1126/science.1109727 ببمد: 15749690
- ^ Koechlin, E. & Hyafil, A. (October 26, 2007). Anterior prefrontal function and the limits of human-decision making. Science, Vol. 318, 594-598.
- ^ Burgess, P.W., Dumontheil, I., & Gilbert, S.J. (2007). The gateway hypothesis of rostral prefrontal cortex (area 10) function. Trends in Cognitive Science, Vol. 11, No. 7.
- ^ Gilbert، Sam J.؛ Spengler، Stephanie؛ Simons، Jon S.؛ Steele، J. Douglas؛ Lawrie، Stephen M.؛ Frith، Christopher D.؛ Burgess، Paul W. (1 يونيو 2006). "Functional specialization within rostral prefrontal cortex (area 10): a meta-analysis". Journal of Cognitive Neuroscience. ج. 18 ع. 6: 932–948. DOI:10.1162/jocn.2006.18.6.932. ISSN:0898-929X. PMID:16839301.